Le but de cette présentation sera d’explorer un nouveau modèle de la physiologie de l’effort et de préciser le fonctionnement des différentes filières énergétiques que l’on connaît déjà.
Cahier des charges
Pour s’assurer qu’on soit tous sur la même page, on va commencer par passer en revue le modèle énergétique populaire.
On parlera ensuite des limitations de ce modèle et notamment des problèmes qu’on trouve quand on compare ce modèle aux observations pratiques avec des outils de mesure comme le Moxy.
Et finalement, on explorera une mise a jour du modèle bioénergétique qui prend en compte différentes études qui sont parues ces 20 dernières années ainsi que les observations pratiques mentionnées au point précédent.
Influences
Tout d’abord il faut mentionner le fait que je n’ai pas créé ce nouveau modèle, j’ai simplement essayé de synthétiser au mieux les informations générées et partagées par des gens beaucoup plus intelligents que moi.
Modèles
Un petit mot sur les modèles avant de commencer.
Les modèles dont on va parler aujourd’hui ne sont pas complets ou exactes – certains aspects sont absents alors que d’autres éléments sont simplifiés pour aider à la compréhension et maintenir une vue d’ensemble.
Vous trouverez certainement aussi des erreures dans mes propos ou dans les idees presentees. Je vous invite donc à partager vos commentaires après avoir regardé cette vidéo afin que l’on puisse tous bénéficier des échanges qui suivront et, je l’espère, mettre à jour nos modèles respectifs.
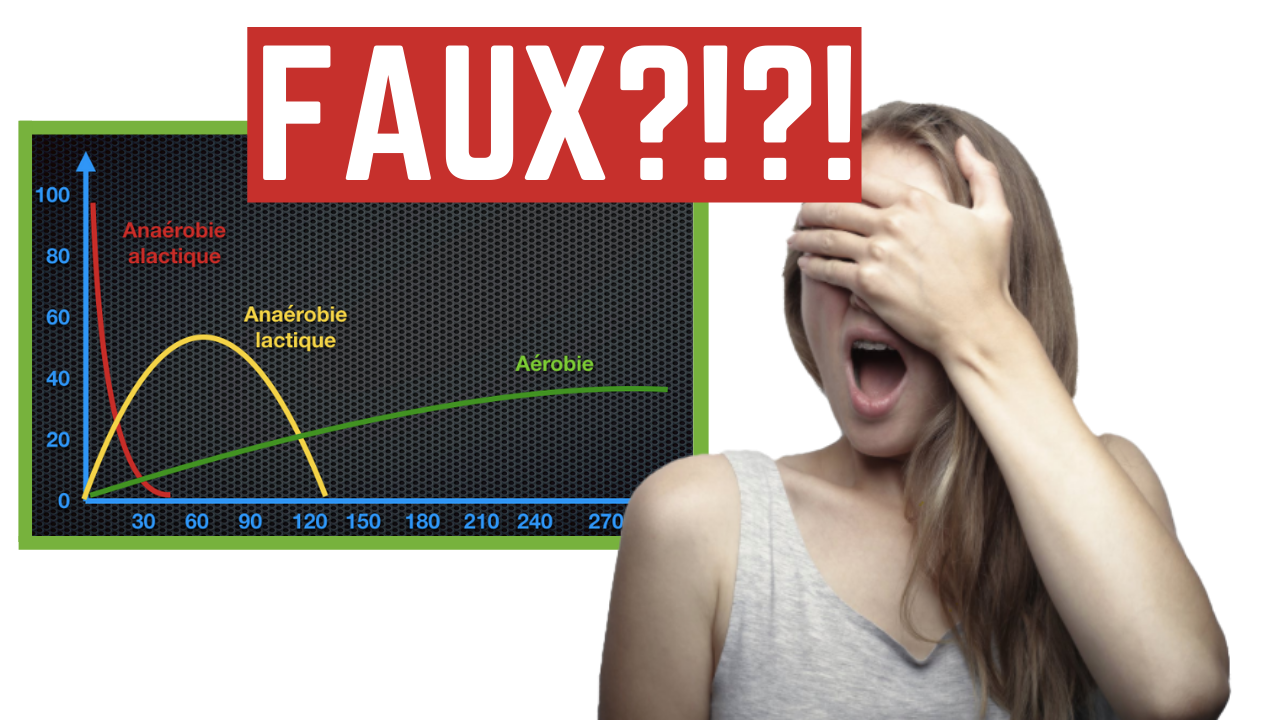
Modèle populaire
Commençons donc avec le modèle bioénergétique que tout le monde connaît.
On sait tout d’abord que tout effort physique requiert de l’ATP, qui est l’unité énergétique du corps humain.
Pour alimenter une contraction musculaire, l’ATP est convertie en ADP, ce qui libère l’énergie contenue dans ses liaisons chimiques. L’ATP doit ensuite être reconstitué afin de permettre de nouvelles contractions musculaires.
C’est la que les filières énergétiques entrent en jeu.
Les filières énergétiques sont des stratégies que l’organisme a développé pour reconstituer les molécules d’ATP nécessaires au bon fonctionnement de l’organisme.
Dans le modèle populaire, on met l’accent sur trois filières énergétiques distinctes.
On parle tout d’abord de la filière anaérobie alactique, filière qui comme son nom l’indique, fonctionne sans oxygène et sans produire de lactate. Cette filière utilise la phosphocréatine pour convertir l’ADP en ATP. Cette filière a la capacité de reconstituer beaucoup d’ATP très rapidement, mais ne peut fonctionner que pour quelques instants. En général, on parle d’une durée de fonctionnement de 3 à 15 secondes pour les efforts intenses.
On a ensuite la filière anaérobie lactique, qui fonctionne également en l’absence d’oxygène. Cette filière est animée principalement par un processus qu’on appelle la glycolyse, qui convertit du glucose (qui est donc le sucre contenu dans le sang) en pyruvate et en lactate (d’où son nom). Cette réaction engendre également la production de molécules d’ATP. On dit que cette filière est la source principale de l’énergie fournie pour des efforts qui durent entre 30 et 120 secondes.
Et finalement, on a la filiere aerobie. Cette filière dégrade des graisses et des sucres avec l’aide de l’oxygène pour générer beaucoup d’ATP. Cependant, on dit souvent que cette filière est lente à se mettre en marche, et intervient principalement sur les efforts moins intenses de moyennes et de longues durées. La mesure phare de catte capacité est la VO2max.
Mais ce modèle présente quelques problèmes.
Limitations du modèle populaire
Premièrement, on sait que les filières glycolytiques et oxydatives n’ont pas la capacite de generer l’ATP assez rapidement pour faire face aux demandes énergétiques des contractions musculaires. Pour donner un exemple concret, la vitesse de consommation d’oxygène par la filière aérobie est environ 1000x plus lente que la vitesse à laquelle la phosphocréatine est utilisée pour recomposer l’ATP dans la filière dite anaérobie alactique.
Deuxièmement, on sait depuis les année 80 avec les travaux de George Brooks que le lactate est présent dans le corps à tout moment, même au repos. Cela va à l’encontre de l’idée que la filière qu’on appelle anaérobie lactique ne fonctionne qu’à partir de 30 secondes d’effort.
Troisièmement, ce modèle populaire n’explique pas le rôle du glycogène dans le processus énergétique de l’effort. Le glycogène qualifie les sucres qui sont stockés dans les muscles et dans le foie.
Et finalement, on a maintenant des technologies qui nous permettent de voir en temps réel le taux d’oxygène présent dans le muscle pendant l’effort. Si on étudie même brièvement les données qui sont fournies par de telles instruments, on voit très vite que l’oxygène est utilisé instantanément, dès le commencement de l’effort. Que ce soit dans le cadre d’un sprint ou d’un marathon, l’oxygène joue un rôle prépondérant dans le processus énergétique.
Utilisation de l’oxygène
Pour illustrer ce dernier point, on peut regarder une étude conduite par McCully et al où ils ont mesurer la saturation d’oxygène dans le muscle avant, pendant, et après une activité physique.
On voit la ligne noire, qui représente le taux d’oxygène, qui descend dès le début de l’activité physique et qui remonte dès que l’activité est interrompue.
Point intéressant à relever: les petites cercles représentent le taux de Phosphocréatine dans le muscle. On voit donc que la le stock de PCr se renouvelle avec une dynamique très similaire à celle du taux d’oxygène présent dans le muscle. On reviendra sur ce point un peu plus tard.
Alors comment pouvons nous réconcilier l’ancien modèle bioénergétique avec ces nouvelles trouvailles?
Glycogen Shunt Model
La piece centrale de ce nouveau modèle bioénergétique vient d’une étude de 2001 intitulée “The “glycogen shunt” in exercising muscle: A role for glycogen in muscle energetics and fatigue.” par Shulman et al.
Ce modèle du “glycogen shunt” ou détournement du glycogène, est un modèle de l’énergétique de la contraction musculaire.
On sait qu’une contraction musculaire prend entre 10 et 40 millisecondes, ce qui signifie que l’énergie nécessaire pour alimenter de nouvelles contractions doit être disponible dans un délai extrêmement court.
Comme on l’a vu précédemment, ce délai est trop court pour que la glycolyse ou la filière aérobie soient des contributeurs directs.
La contraction musculaire est alimentée par l’énergie contenue dans les liaisons chimiques de l’ATP. En raison des fortes demandes soudaines d’énergie, seule la voie dite anaérobie alactique (fonctionnant avec la phosphocréatine) peut fournir suffisamment d’ATP pour alimenter ce processus.
La phosphocréatine «donne» alors une molécule de phosphate à l’ADP pour former à nouveau l’ATP. Cela se produit dans les 15 premières millisecondes suivant le début de la contraction musculaire.
Dans les ≈85 millisecondes suivantes, la glycogénolyse décompose le glycogène en lactate, ce qui fournit l’ATP nécessaire pour resynthétiser la phosphocréatine et reconstituer le stock appauvri. Cela permet ainsi de continuer à alimenter les prochaines contractions musculaires. C’est ce qu’on appelle le “glycogen shunt”.
Alors que le glycogène est décomposé pour synthétiser l’ATP, le glycogène est simultanément re-synthétisé de manière à maintenir des niveaux de glycogène suffisant dans le muscle. L’ATP nécessaire pour alimenter ce processus est fourni par l’oxydation du lactate.
Tant que des niveaux suffisants d’oxygène sont disponibles dans le muscle, l’oxydation du lactate fournit l’ATP nécessaire pour resynthétiser le glycogène et permet ainsi à la glycogénolyse d’être un contributeur majeur au processus énergétique.
Mais quand l’oxygène vient à manquer, le glycogène reste constant, et le «vide» énergétique est comblé par une régulation à la hausse de la glycolyse. Et vu que la glycolyse est moins rapide que la glycogénolyse, l’intensité de l’effort est inévitablement réduite.
Ce modèle pourrait également expliquer le rôle du glycogène dans la fatigue pendant l’exercice, où de faibles concentrations de glycogène ne permettent pas à la voie de glycogénolyse de fournir l’ATP nécessaire pour resynthétiser la phosphocréatine pendant une contraction musculaire.
Glycogen Shunt en bref
Pour résumer, l’ATP qui est nécessaire pour recomposer les molécules de phosphocréatine est générée via la filière de la glycogénolyse. Cette dernière a la capacité de subvenir aux hautes demandes énergétiques de la filière qu’on appelle anaérobie alactique.
Tant que l’oxygène est disponible en quantité suffisante, le stock de glycogène est recomposé de manière continue avec l’aide de la filière aérobie.
Mais lorsque le taux d’oxygène est réduit, la filière de la glycolyse, qu’on appelait avant anaérobie lactique, contribue plus fortement au recyclage de la phosphocréatine.
Tous ces processus sont actifs quasi instantanément dès le début d’une contraction musculaire.
Mise à jour
A l’aide de ces nouvelles informations on peut maintenant mettre à jour notre modèle des filières énergétiques.
Plutôt que d’avoir trois filières qui travaillent de manière indépendantes, on a maintenant 4 filières qui travaillent les unes avec les autres dès le premier instant d’effort.
Ce qui est intéressant avec ce nouveau modèle c’est qu’il est applicable à toutes les disciplines sportives, du sprint au marathon en passant par les sports collectifs. On verra simplement des variations entre chaque sport notamment au niveau de la vitesse à laquelle l’oxygène est utilisé, des seuils maximum et minimum de saturation d’oxygène dans le muscle, et de la contribution relative de la glycolyse au processus énergétique global.
Considerations pratiques
On voit donc que l’oxygène joue un rôle central à la performance, peu importe la durée de l’effort.
On peut aussi en déduire que pour améliorer les performances sportives, il faut optimiser l’approvisionnement et l’utilisation de l’oxygène.
Un sprinter doit être capable par exemple d’utiliser tout l’oxygène disponible très rapidement pour alimenter un effort très intense, alors qu’un marathonien devra favoriser un approvisionnement d’oxygène pour une durée prolongée, sans jamais complètement vider son réservoire d’oxygène.
Modèle des systèmes limitants
Cela nous amène vers un autre modèle de performance sportive présenté par Juerg Feldmann il y a déjà plusieurs années. Juerg parle de l’idée des systèmes qui sont impliqués dans la performance sportive, systèmes qui, une fois qu’ils arrivent à leur capacité maximale, vont limiter nos performances et induire des compensations.
Ces systèmes sont au nombre de 4:
Il y a tout d’abord le cerveau qui joue le rôle de gouverneur central et module les impulsions au niveau du système nerveux.
On a ensuite le système respiratoire qui permet de capturer l’oxygène dans l’air ainsi que de se débarrasser du Co2 produit lors du processus aérobie.
Le coeur et le système circulatoire distribuent l’oxygène et permettent aussi de déplacent d’autres molécules dans le corps comme le lactate et le CO2.
Et finalement, on a les tissus musculaires qui eux utilisent l’oxygène pour alimenter les contractions musculaires.
Plus le sport est intense et de courte durée (comme l’haltérophilie ou le powerlifting par exemple), plus le système nerveux est impliqué et peut devenir un facteur qui limitera la performance. Pour les disciplines de plus longue durée, c’est les systèmes respiratoire, circulatoire et musculaire qui viendront généralement limiter les performances.
Une fois qu’un système atteint sa limite de contribution au processus énergétique, les autres systèmes vont essayer de compenser ce déficit. Mais ils ne pourront le faire que pour une durée limitée avant que l’intensité de l’effort ne doive être réduite.
L’étape suivante
A l’aide de certains protocoles de test, on peut déterminer quel(s) système(s) limite la performance d’un athlète, et ainsi véritablement individualiser le programme d’entraînement en se focalisant sur le ou les systèmes limitants.
Si vous êtes intéressés à en apprendre plus sur les protocoles de test, les outils qui sont impliqués, l’interprétation des résultats et la créations de programmes d’entraînements adaptés à ce nouveau paradigme, je vous invite à suivre et à écouter les leaders en la matière dans le monde de la performance. Vous trouverez les liens et des ressources dans la description de la vidéo.
Merci d’avoir pris le temps de regarder cette présentation! J’attends vos commentaires avec impatience, et n’oubliez pas de liker la vidéo, de la partager et de vous abonner a la chain. A bientot!
Études citées:
McCully et al (1994). Simultaneous in Vivo Measurements of HbO2 Saturation and PCr Kinetics After Exercise in Normal Humans. Journal of Applied Physiology
https://pubmed.ncbi.nlm.nih.gov/7961273/ (02.06.2020)
Shulman, R. G., & Rothman, D. L. (2001). The “glycogen shunt” in exercising muscle: A role for glycogen in muscle energetics and fatigue. Proceedings of the National Academy of Sciences of the United States of America. https://doi.org/10.1073/pnas.98.2.457 (17.05.2020)
Autres resources:
George A. Brooks (2020). Lactate as a fulcrum of metabolism. Exercise Physiology Laboratory, Department of Integrative Biology, University of Caroline, Berkeley, CA
https://www.sciencedirect.com/science/article/pii/S2213231720300422 (02.06.2020)
Drouin et al (2019). Fatigue-independent Alterations in Muscle Activation and Effort Perception During Forearm Exercise: Role of Local Oxygen Delivery. Journal of Applied Physiology. https://pubmed.ncbi.nlm.nih.gov/31070953/ (03.06.2020)
Jensen et al (2011). The Role of Skeletal Muscle Glycogen Breakdown for Regulation of Insulin Sensitivity by Exercise. Frontiers in Physiology.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3248697/ (02.06.2020)
Chung et al (1998) Metabolic Fluctuation During a Muscle Contraction Cycle. The American Journal of Physiology.
https://pubmed.ncbi.nlm.nih.gov/9530118/ (02.06.2020)
Llavero et al (2019) McArdle Disease: New Insights into Its Underlying Molecular Mechanisms. International Journal of Molecular Sciences.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6929006/ (02.06.2020)
Evan Peikon:
Evan sur Instagram: https://www.instagram.com/evan_peikon/
Evan sur Youtube: https://www.youtube.com/channel/UCVGrO03fD_TdUX_6ak3vNvA
Interview avec Evan: https://youtu.be/svBYe_h61ok
Aaron Davis:
Aaron sur Instagram: https://www.instagram.com/davisxctf/
Interview avec Aaron: https://youtu.be/BKeVSMw-8C0
Evolve Health & Performance website: https://www.evolvehp.life/
Evolve Health & Performance sur Instagram: https://www.instagram.com/evolve_hpl/
Justin Moore:
Justin on Instagram: https://www.instagram.com/jmsb_strengthtraining/
Justin’s blog: https://justinmoore.home.blog/
Moxy (outil de mesure):
Webinars gratuits: https://www.youtube.com/user/MoxyMonitor
Andri Feldmann:
Andri sur Youtube: https://www.youtube.com/channel/UC24U0DXcMIHpgvWT0zE6E8g